【過酸化水素による細胞の形態変化と老化関連βガラクトシダーゼ活性抑制作用】
過酸化水素に暴露された細胞は加齢早期の形態変化を起こす。さらに、老化関連βガラクトシダーゼ活性が増加することが報告されている。冠元穎粒は老化に関係する細胞の形態変化を抑制し、老化関連βガラクトシダーゼ活性も阻害した(図6)。
図6.H202によるSA-8-Gal活性
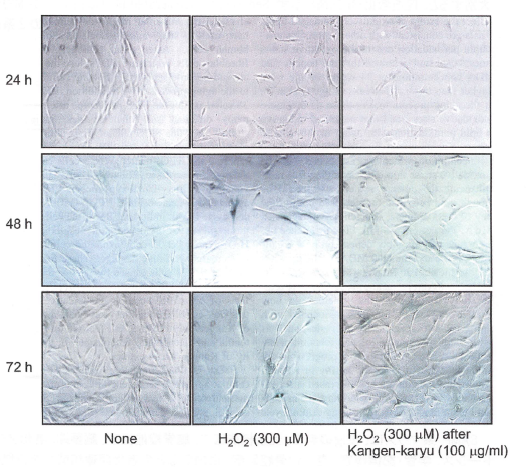
ヒト線維芽細胞は培地か冠元穎粒(100″g/mL)存在下で24時間培養し、60分間過酸化水素で処置された。過酸化水素除去後、細胞は24,48および72時間新鮮な培地で培養し、老化関連βガラクトシダーゼ染色と細胞の形態観察を行った(倍率40倍)。
【還元穎粒の細胞増殖に対する効果】
過酸化水素の酸化的ストレスにより細胞増殖は抑制された。還元穎粒の前処置は酸化的ダメージにより生じた増殖抑制を減弱した(図7)。
図7.冠元穎粒の細胞増殖に対する影響
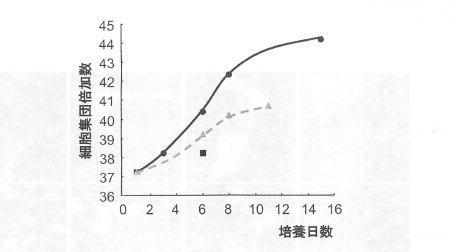
過酸化水素(300uM)をヒト線維芽細胞に添加し、細胞の倍加数を測定した。
●は無処置群、■は過酸化水素処置群、△は過酸化水素+100ug/mL冠元穎
粒処置群を示す。
【冠元穎粒の転写因子NF-“B核内移行に対する影響】
無刺激細胞ではNF-“Bは細胞質に存在する(図8,None)。過酸化水素の添加はNF-“Bの核内への移行を促進した(図8、H202)。冠元穎粒の前処置は、過酸化水素によるNF-凡Bの核内移行を濃度依存的に抑制した(図8、H202 (300MM)afterKangen-karyu(10,100ug/mL))。
図8. NF-“Bのトランスロケーション.
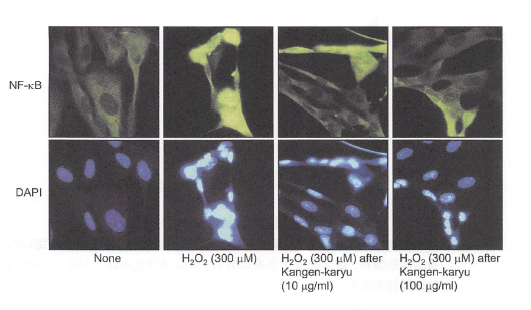
ヒト線維芽細胞を冠元穎粒(10, 100ug/mL)で24時間前処置し、その後、過酸化水素(300MM)を60分間添加した。細胞はNF-“B抗体で処置し、それからDAPI染色した。NF-“Bの核への転移は蛍光顕微鏡で観察した(倍率X100)。
【冠元穎粒のミトコンドリア膜電位への影響】
ローダミンー123はミトコンドリアの膜電位の減少に伴い、ミトコンドリア内に取り込まれることが知られている。図9に見られるように、過酸化水素で刺激したヒト線維芽細胞は、無刺激の細胞と比較して明らかにローダミン123による蛍光強度の増加が観察された。冠元穎粒の前処置は、膜電位の減少を抑制した(図9)。
図9.ミトコンドリアの膜電位に対する冠元穎粒の影響.
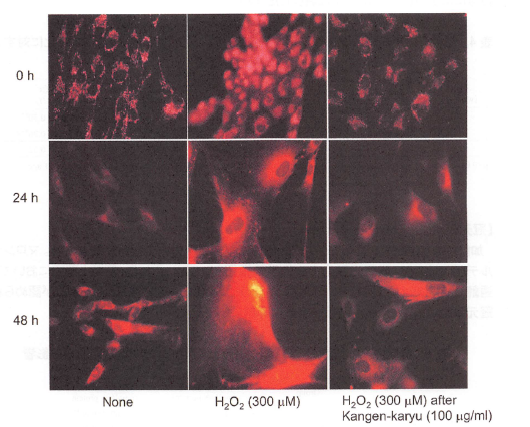
ヒト線維芽細胞に冠元穎粒(100ug/mL)を24時間前処置し、次に60分間過酸化水素を加え、その後細胞を0,24,48時間培養した。ミトコンドリア膜電位はローダミン123染色により赤色蛍光を呈する細胞を蛍光顕微鏡下で観察した(倍率xl00倍)。
【老化促進ラット(SAMP)における冠元穎粒の抗酸化作用】
*冠元穎粒による.OH産生抑制
メチルグアニンジン(MG)はヒドロキシラジカルによってクレアチニン(Cr)より合成される。
MG/Cr比はヒドロキシラジカル産生の指標である。34週齢のSAMPでは、コントロール(5週齢SAMP)と比較して有意な尿中MG、C「、MG/Crの上昇が見られた。一方、冠元穎粒投与によりMG/Cr値は有意に減少した(表4)。
表4。尿中クレアチニン、メチルグアニジンとメチルグアニジン/クレアチニン比に対する冠
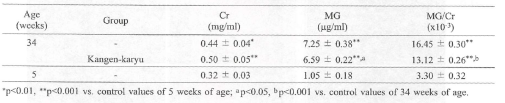
【冠元穎粒による脂質過酸化抑制】
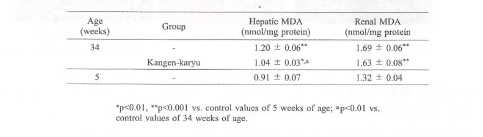
加齢により細胞内の脂質、タンパクそしてDNAの酸化物の蓄積が増加する。マロンジアルデヒド(MDA)は脂質過酸化の生物学的指標として用いられる。肝臓と腎臓において34週齢SAMPではコントロール(5週齢SAMP)と比較して、有意なMDAの増加が認められた。
冠元穎粒は肝臓のMDA濃度を有意に抑制した(表5)。
表5.肝臓および腎臓のマロンジアルデヒド濃度に対する冠元穎粒の影響
【冠元穎粒によるタンパク酸化抑制】
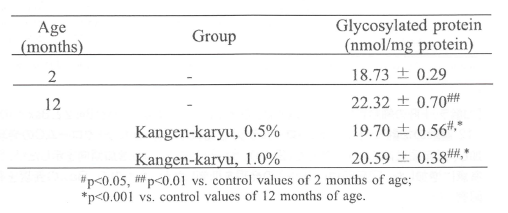
加齢とともに体内では最終糖化生成物が蓄積することが知られている(繁田幸男ら、蛋白の糖化AGEの基礎と臨床、医学書院(1997) 、今泉勉、AGEs研究の最前線、メディカルレビュー社(2004)。このAGEsは、糖と蛋白質が非酵素的に結合する糖化(メイラード反応)により生成される。表6に示すように、コントロール(2週齢SAMP)と比較して加齢ラット(12週齢SAMP)では、血清中糖化タンパクの有意な上昇を認めた。冠元穎粒は加齢により増加した糖化タンパクの生成を抑制した。
表6.血清中糖化タンパク濃度に対する冠元穎粒の影響
【冠元穎粒によるグルタチオン/酸化型グルタチオン比(GSH/GSSG)への影響】
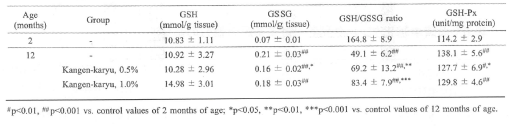
グルタチオン(GSH)は強い抗酸化物質である。グルタチオンペルオキシダーゼ(GSH-Px)がリン脂質の酸化を抑制することにより、生体内のGSHは酸化型グルタチオン(GSSG)に変化することが知られている。加齢は、肝臓組織においてGSH濃度の変化を伴うことなくGSSG量を増加させることが報告されている。表7に示すように、12週齢の加齢ラットではGSSGの増加とGSH/GSSG比の低下が認められたが、冠元穎粒投与によりGSSG濃度は低下し、GSH/GSSG比は上昇した。一方GSH-Px活性は加齢と共に促進され、冠元穎粒により有意な低下を認めた(表7)。
表7.冠元穎粒のGSH、GSSG、GSH/GSSG比およびGSH-Px活性に対する影響
【加齢ラット肝の細胞質チトクロームcおよびアポトーシス関連タンパクBcI-2とBaxへの影響】
12週齢の加齢ラットではコントロールラットと比較して明らかにチトクロームCの発現が増加し、冠元穎粒はこれを抑制した(図10)。BcI-2は加齢により増加傾向を示したが、Baxは有意に増加した。冠元穎粒はBcI-2の発現を抑制しなかったものの、Baxの発現を有意に阻害した。
図10.細胞質のチトクロームc、BcI-2そしてBaxに対する冠元穎粒の影響
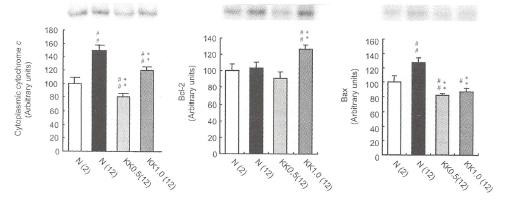
各タンパクの発現はウェスタンプロット法にて検出された。#p<0.01、##p<0.001 vs。2週齢コントロ ールラット値、*p<0.05、**p<0.001vs.12週齢ラットのコントロール値と比較したときの有意差。 【炎症に関わる因子に対する冠元穎粒の影響】 活性酸素は、前炎症性プロスタノイドの合成に関わるシクロオキシゲナーゼー2(COX-2)発現を促進する。また、誘導型一酸化窒素合成酵素(iNOS)やヘムオキシゲナーゼ(HO-1)も活性酸素産生に依存して炎症性サイトカインの誘導に関与する。加齢ラットにおいてCOX-2、iNOSおよびHO-1の発現を促進した。冠元穎粒はこれらの促進を抑制した(図11)。 図11.加齢ラットのCOX-1、COX-2、iNOSおよびHO-'発現に対する冠元穎粒の影響
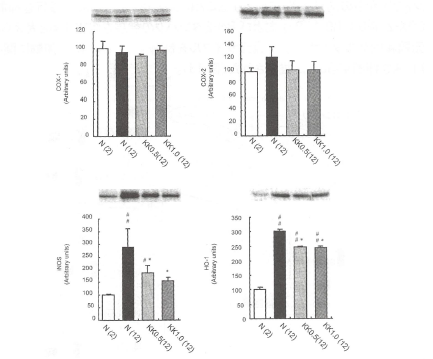
タンパクの発現量は濃度測定機で測定した。#p<0.01,##0.001vs.,controlvalue of 2 monthsofageand*p<0.001vs。controlvalueofl2months.